
Part:BBa_K1998007
psbMZHWK-OPQR
Sequence and Features
- 10COMPATIBLE WITH RFC[10]
- 12COMPATIBLE WITH RFC[12]
- 21INCOMPATIBLE WITH RFC[21]Illegal BglII site found at 3063
Illegal BglII site found at 3392 - 23COMPATIBLE WITH RFC[23]
- 25INCOMPATIBLE WITH RFC[25]Illegal NgoMIV site found at 570
Illegal NgoMIV site found at 1819
Illegal NgoMIV site found at 2465 - 1000INCOMPATIBLE WITH RFC[1000]Illegal BsaI site found at 3456
Overview
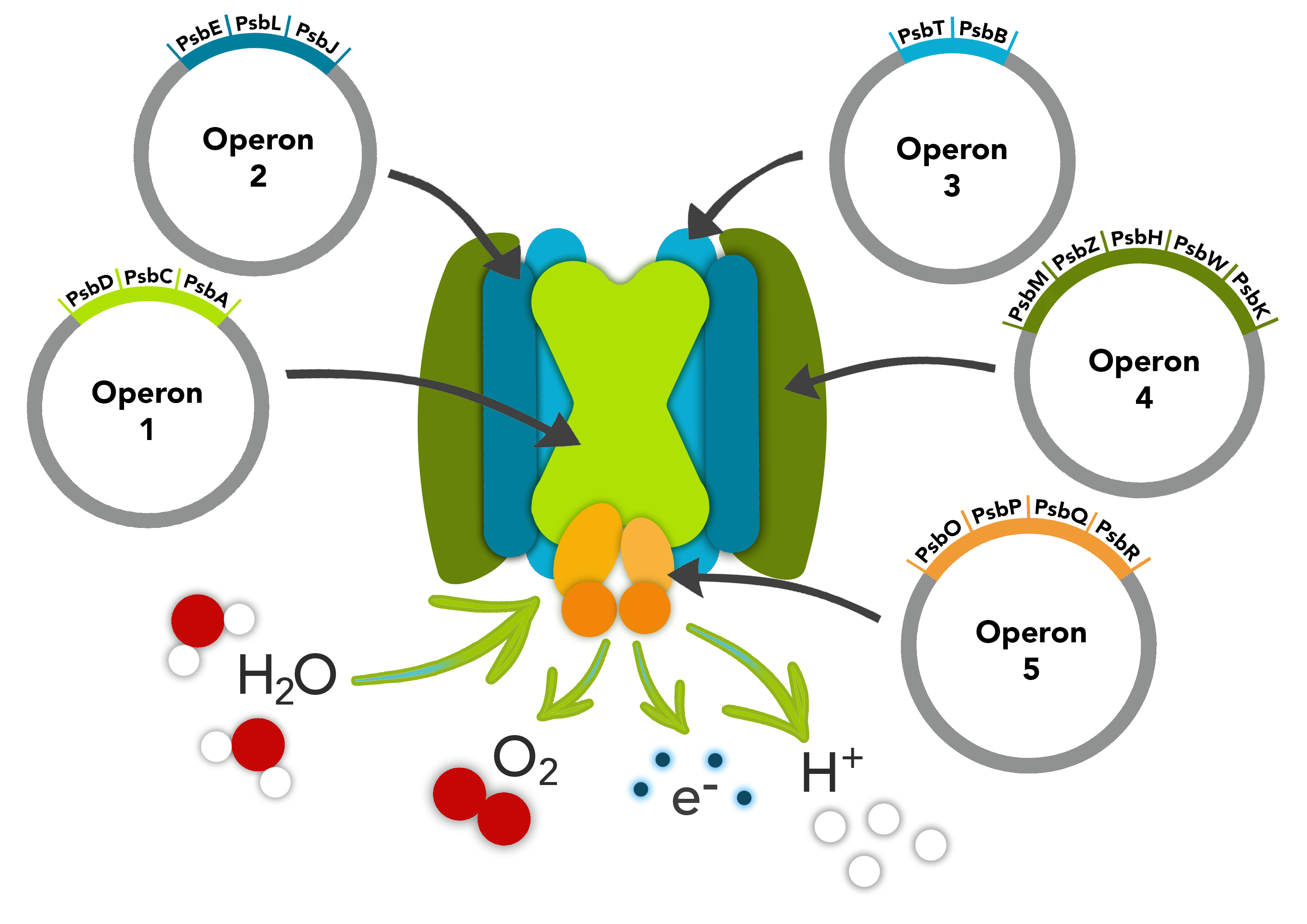
This part is composed of two operons in the photosystem II pathway; psbMZHWK and psbOPQR. The psbM, psbZ, psbH, psbW and psbK compose the first operon. The psbM protein subunit is positioned at the monomer-monomer interface. The psbZ protein controls the interaction of Photosystem II cores with the light-harvesting antenna. The psbH protein is required for stability and assembly of the photosystem II complex. The psbW protein stabilizes dimeric photosytem II. The psbK protein is also required for stability and assembly of Photosystem II. The second operon is composed of the psbO, psbP, psbQ and psbR genes. The psbO gene that comprises this part codes a manganese stabilizing protein. The psbP gene that comprises this part transcribes a protein that optimizes the availability of Ca2+ and Cl- cofactors in the Oxygen Evolving Complex in PSII to maintain the active Manganese cluster. The psbQ protein is an oxygen enhancer protein. The psbR protein is an important link in the Photosystem II core complex for stable assembly of the Oxygen Evolving Complex.
These parts make up two of the operons in our PSII pathway.
Biology & Literature
The first gene in this operon is psbM which acts to stabalise the dimerisation of the PSII complex through it's span over the membrane by way on an α-helix located at the monomer-monomer interface of PSII [1]. It has been shown that it's absence will weaker the dimer interconnection of the core complex and may impair PSII repair, however it is not a necessary factor for PSII biosynthesis [2]. In addition the success of PSII centre assembly decreases when this gene is removed [3].
The psbZ gene is also referred to as ycf9 [4]. The gene highly conserved gene amongst photosynthetic species and interacts with the light harvesting antenna in the PSII complex [4], found close to the PSII interface and light harvesting complex II [5]. The two transmembrane helix protein which it encodes [1], has been indicated to result in decreased stability of the both the PSII and Light Harvesting Complex II if deleted [4]. This suggests that it has a role in anchoring these two complexes.
The third gene in this operon, psbH contains a membrane spanning alpha helix [6] and contains multiple phosphorylation sites [7]. Deletion of the psbH complex destabilises the PSII complex and affects the ability of bicarbonate to bind to the complex [8]. In another studying the elimination of the gene in Chlamydomonas reinhardtii, affects the ability of the PSII complex to form, therefore indicating the importance of the psbH gene for the synthesis of Photosystem II [9].
The next subunit protein in the operon is encoded by the psbW gene which is a transmembrane protein, responsible for photo-protection and dimer stabilisation [10]. It has been shown to have a close association with the PSII reaction centre [11]. The absence of psbW affects the formation of the PSII complex [12, 13] and it's absence in transgenic plants have indicated a higher sensitivity to light based stress [14].
The fifth gene in this operon encodes psbK, another subunit of PSII involved in maintaining the stability of the complex [15]. It has been shown to be tightly associated with the PSII complex, in particular the CP43 antenna complex [16]. Some destabilisation of the PSII complex can be detected if removed from C. reinhardtii [15] however it is not necessary for the formation of the complex.
As part of the next series of genes, the gene, psbO in this operon is attached to the luminal membrane of PSII [17] and through a cluster of Mn2+ forms the centre of the oxygen evolving complex [18]. Described as having an eight strand β-barrel, a large loop between strands five and six connects the oxygen evolving complex to the luminal surface [19]. Inactivation of the psbO gene in Chlamydomonas reinhardtii prevents the assembly of PSII [20]. In Synechocystis sp. however, deletion of the psbO gene has not had a major effect on oxygen evolution, but a larger effect on the susceptibility of photo inhibition can be seen [21]. Therefore it can be concluded that the whilst the psbO gene is not essential to the assembly of the PSII complex or the water-splitting pathway it is important in providing protection to the PSII from damage caused by light.
The psbP gene is one of the extrinsic proteins found in higher plants and algae [22]. It has a two domain structure made up of an antiparallel b-sheet and a b-strand that form domain 1. Domain 1 lies back to back with a central b-sheet (domain II) [23]. Decrease oxygen evolving activity was detected when psbP has not been present in C. reinhardtii [20]. The 23kDa protein is involved in PSII water oxidation process [24]. The psbP gene that comprises this part transcribes a protein that optimizes the availability of Ca2+ and Cl- cofactors in the Oxygen Evolving Complex in PSII to maintain the active Manganese cluster [25].
The psbQ gene in C. reinhardtii binds directly the PSII complex independent of other extrinsic proteins [26]. The protein is found on the inner side of the thylakoid lumen with a polyproline II helix [27]. It helps aid PSII to function in conditions where there is low light [28]. Further to this, absence of psbQ has indicated an impairment of the ability of cells to generate oxygen as well as impacting on the presence of psbV destabilised [29].
The final gene in this operon is the psbR. This gene encodes a 10kDa protein and whilst the genes role is not certain, it is understood that the gene is necessary for the optimisation of electron transfer and water oxidation [30] as the absence of psbR causes instability in the PSII complex. Other psbR mutants have indicated similar decrease in oxygen evolution [31, 32].
Part Verification

Fig 1. Gel electrophoresis of the operons and single parts constituting the Photosystem II pathway implemented in this project. The part psbMZHWK-OPQR (3653 bp) can be seen in lane 9 on the gel at the correct size relative to the 1kb ladder and other parts in our project shown on this gel. The band lower down in this lane on the gel is the pSB1C3 backbone (2000 bp).
Protein information
psbM
mass: 3.76kDa
sequence: MEVNIYGLTATALFIIIPTSFLLILYVKTASTQD
psbZ
mass: 4.56kDa
sequence: MVGVPVVFATPNGWTDNKGAVFSGLSLWLLLVFVVGILNSFVV
psbH
mass: 6.02kDa
sequence: MSEAGKVLPGWGTTVLMAVFILLFAAFLLIILEIYNSSLILDDVSMSWETLAKVS
psbW
mass: 9.2kDa
sequence: MATTVRSEVAKKVAMLSTLPATLAAHPAFALVDERMNGDGTGRPFGVNDPVLGWVLLGVFGTMWAIWFIGQKDLGDFEDADDGLKL
psbK
mass: 5.0kDa
sequence: MTTLALVLAKLPEAYAPFAPIVDVLPVIPVFFILLAFVWQAAVSFR
psbO
mass: 27.96kDa
sequence:
MAQKVGQAAAAAALATAMVAGSANALTFDEIQGLTYLQVKGSGIANTCPVLESGTTNLKELKAGSYKLENFC IEPTSFTVKEESQFKGGETEFVKTKLMTRLTYTLDAMSGSFKVGSDGSAELKEDDGIDYAATTVQLPGGERV AFLFTIKQFDGKGTLDNIKGDFLVPSYRGSSFLDPKGRGGSTGYDNAVALPARADAEELLKENVKITKALKG
SAVFSVAKVDPVTGEIAGVFESIQPSDTDLGAKPPKDIKVTGLWYAQLK*
psbP
mass: 23.17kDa
sequence:
MASGSDVSRRAALAGFAGAAALVSSSPANAAYGDSANVFGKVTNKSGFVPYAGDGFALLLPAKWNPSKENDF PGVILRYEDNFDAVNNLVVIAQDTDKKAIADFGSQDKFLESVSYLLGKQAYSGETQSEGGFAPNRVSAASLL DVSTTTDKKGKTYYKYELLVRSADGDEGGRHQLIGATVGSDNKLYIIKIQIGDKRWFKGAKKEAMGAFDSFTVV*
psbQ
mass: 19.6kDa
sequence:
MASGESRRAVLGGLLASAVAAVAPKAALALTPVDLFDDRSVRDRGFDLIYEARDLDLPQNVREGFTQARASL
DETKKRVKESEARIDADLDVFIQKSYWTEAREQLRRQVGTLRFDLNTLASTKEKEAKKAALGLRKEFIQAVED
LDFALREKDQASAAKKLEITKAKLDSVLAAVL
psbR
mass: 12.2kDa
sequence:
MGGGKTDITKVGLNSIEDPVVKQNLMGKSRFMNKKDWKDASGRK
References
[1] Ferreira KN, Iverson TM, Maghlaoui K, Barber J, Iwata S. Architecture of the photosynthetic oxygen-evolving center. Science. 2004 Mar 19;303(5665):1831-8.
[2] Umate P, Schwenkert S, Karbat I, Dal Bosco C, Mlcòchová L, Volz S, Zer H, Herrmann RG, Ohad I, Meurer J. Deletion of psbM in tobacco alters the QB site properties and the electron flow within photosystem II. Journal of Biological Chemistry. 2007 Mar 30;282(13):9758-67.
[3] Bentley FK, Luo H, Dilbeck P, Burnap RL, Eaton-Rye JJ. Effects of Inactivating psbM and psbT on Photodamage and Assembly of Photosystem II in Synechocystis sp. PCC 6803†. Biochemistry. 2008 Oct 4;47(44):11637-46.
[4] Swiatek M, Kuras R, Sokolenko A, Higgs D, Olive J, Cinque G, Müller B, Eichacker LA, Stern DB, Bassi R, Herrmann RG. The chloroplast gene ycf9 encodes a photosystem II (PSII) core subunit, psbZ, that participates in PSII supramolecular architecture. The Plant Cell. 2001 Jun 1;13(6):1347-68.
[5] Minagawa J, Takahashi Y. Structure, function and assembly of Photosystem II and its light-harvesting proteins. Photosynthesis research. 2004 Dec 1;82(3):241-63.
[6] Michel HP, Bennett J. Identification of the phosphorylation site of an 8.3 kDa protein from photosystem II of spinach. FEBS letters. 1987 Feb 9;212(1):103-8.
[7] Vener AV, Harms A, Sussman MR, Vierstra RD. Mass spectrometric resolution of reversible protein phosphorylation in photosynthetic membranes ofArabidopsis thaliana. Journal of Biological Chemistry. 2001 Mar 9;276(10):6959-66.
[8] Komenda J, Lupinkova L, Kopecký J. Absence of the psbH gene product destabilizes photosystem II complex and bicarbonate binding on its acceptor side in Synechocystis PCC 6803. European Journal of Biochemistry. 2002 Jan 1;269(2):610-9.
[9] Summer EJ, Schmid VH, Bruns BU, Schmidt GW. Requirement for the H phosphoprotein in photosystem II of Chlamydomonas reinhardtii. Plant physiology. 1997 Apr 1;113(4):1359-68.
[10] Woolhead CA, Mant A, Kim SJ, Robinson C, Rodger A. Conformation of a purified “spontaneously” inserting thylakoid membrane protein precursor in aqueous solvent and detergent micelles. Journal of Biological Chemistry. 2001 May 4;276(18):14607-13.
[11] Shi LX, Schröder WP. Compositional and topological studies of the psbW protein in spinach thylakoid membrane. Photosynthesis research. 1997 Jul 1;53(1):45-53.
[12] Shi LX, Lorković ZJ, Oelmüller R, Schröder WP. The low molecular mass psbW protein is involved in the stabilization of the dimeric photosystem II complex in Arabidopsis thaliana. Journal of Biological Chemistry. 2000 Dec 1;275(48):37945-50.
[13] García‐Cerdán JG, Kovács L, Tóth T, Kereïche S, Aseeva E, Boekema EJ, Mamedov F, Funk C, Schröder WP. The psbW protein stabilizes the supramolecular organization of photosystem II in higher plants. The Plant Journal. 2011 Feb 1;65(3):368-81.
[14] Thidholm E, Shi LX, Schroder W. The psbW-protein; Its location and involvement in photoinhibition. Science Access. 2001;3(1).
[15] Takahashi Y, Matsumoto H, Goldschmidt-Clermont M, Rochaix JD. Directed disruption of the Chlamydomonas chloroplast <i>psbK gene destabilizes the photosystem II reaction center complex. Plant molecular biology. 1994 Mar 1;24(5):779-88.
[16] Sugimoto I, Takahashi Y. Evidence that the psbK polypeptide is associated with the photosystem II core antenna complex CP43. Journal of Biological Chemistry. 2003 Nov 7;278(45):45004-10.
[17] De Las Rivas J, Barber J. Analysis of the structure of the psbO protein and its implications. Photosynthesis research. 2004 Sep 1;81(3):329-43.
[18} Murata N, Miyao M. Extrinsic membrane proteins in the photosynthetic oxygen-evolving complex. Trends in Biochemical Sciences. 1985 Mar 1;10(3):122-4.
[19] Ferreira KN, Iverson TM, Maghlaoui K, Barber J, Iwata S. Architecture of the photosynthetic oxygen-evolving center. Science. 2004 Mar 19;303(5665):1831-8.
[20] Mayfield SP, Rahire M, Frank G, Zuber H, Rochaix JD. Expression of the nuclear gene encoding oxygen-evolving enhancer protein 2 is required for high levels of photosynthetic oxygen evolution in Chlamydomonas reinhardtii. Proceedings of the National Academy of Sciences. 1987 Feb 1;84(3):749-53.
[21] Mayes SR, Cook KM, Self SJ, Zhang Z, Barber J. Deletion of the gene encoding the Photosystem II 33 kDa protein from Synechocystis sp. PCC 6803 does not inactivate water-splitting but increases vulnerability to photoinhibition. Biochimica et Biophysica Acta (BBA)-Bioenergetics. 1991 Sep 27;1060(1):1-2.
[22] Ifuku K, Yamamoto Y, Ono TA, Ishihara S, Sato F. psbP protein, but not psbQ protein, is essential for the regulation and stabilization of photosystem II in higher plants. Plant Physiology. 2005 Nov 1;139(3):1175-84.
[23] Kuwabara T, Suzuki K. Reversible changes in conformation of the 23-kDa protein of photosystem II and their relationship to the susceptibility of the protein to a proteinase from photosystem II membranes. Plant and cell physiology. 1995 Apr 1;36(3):495-504.
[24] Rova M, Franzén LG, Fredriksson PO, Styring S. Photosystem II in a mutant of Chlamydomonas reinhardtii lacking the 23 kDa psbP protein shows increased sensitivity to photoinhibition in the absence of chloride. Photosynthesis research. 1994 Jan 1;39(1):75-83.
[25] Ido K, Ifuku K, Yamamoto Y, Ishihara S, Murakami A, Takabe K, Miyake C, Sato F. Knockdown of the psbP protein does not prevent assembly of the dimeric PSII core complex but impairs accumulation of photosystem II supercomplexes in tobacco. Biochimica et Biophysica Acta (BBA)-Bioenergetics. 2009 Jul 31;1787(7):873-81.
[26] Suzuki T, Minagawa J, Tomo T, Sonoike K, Ohta H, Enami I. Binding and functional properties of the extrinsic proteins in oxygen-evolving photosystem II particle from a green alga, Chlamydomonas reinhardtii having his-tagged CP47. Plant and cell physiology. 2003 Jan 15;44(1):76-84.
[27] Balsera M, Arellano JB, Revuelta JL, De las Rivas J, Hermoso JA. The 1.49 Å resolution crystal structure of PsbQ from photosystem II of Spinacia oleracea reveals a PPII structure in the N-terminal region. Journal of molecular biology. 2005 Jul 29;350(5):1051-60.
[28] Yi X, Hargett SR, Frankel LK, Bricker TM. The PsbQ protein is required in Arabidopsis for photosystem II assembly/stability and photoautotrophy under low light conditions. Journal of Biological Chemistry. 2006 Sep 8;281(36):26260-7.
[29] Kashino Y, Inoue-Kashino N, Roose JL, Pakrasi HB. Absence of the PsbQ protein results in destabilization of the psbV protein and decreased oxygen evolution activity in cyanobacterial photosystem II. Journal of Biological Chemistry. 2006 Jul 28;281(30):20834-41.
[30] Vinyard DJ, Ananyev GM, Charles Dismukes G. Photosystem II: the reaction center of oxygenic photosynthesis*. Annual review of biochemistry. 2013 Jun 2;82:577-606.
[31] Stockhaus J, Höfer M, Renger G, Westhoff P, Wydrzynski T, Willmitzer L. Anti-sense RNA efficiently inhibits formation of the 10 kd polypeptide of photosystem II in transgenic potato plants: analysis of the role of the 10 kd protein. The EMBO journal. 1990 Sep;9(9):3013.
[32] Suorsa M, Sirpiö S, Allahverdiyeva Y, Paakkarinen V, Mamedov F, Styring S, Aro EM. psbR, a missing link in the assembly of the oxygen-evolving complex of plant photosystem II. Journal of Biological Chemistry. 2006 Jan 6;281(1):145-50.
| None |